
[필립 볼] 기묘한 진화의 필연성(2/2)
단백질 역시 이런 성질을 가지고 있습니다. 단백질중에는 같은 형태로 같은 효소 기능을 하면서도(표현형), 공통된 아미노산은 20% 이하인 단백질들이 있습니다. 매릴랜드에 위치한 국립보건연구원(NIH)의 데이비드 립맨과 W. 존 윌버는 1991년 왜 이런 현상이 발생하는지를 보였습니다. 이들은 단백질의 단순한 모델이 서열 공간의 확장된 네트웍 상에서 중립 돌연변이를 통해 공간을 옮겨 다닐 수 있음을 발견했습니다. 2001년 하버드 대학의 앤터니 키피와 잭 쇼스택은 같은 표현형을 가진 무수히 많은 단백질 서열이 있음을 실험으로 보였습니다. 그들은 임의로 아미노산을 연결한 단백질이 세포에서 에너지 저장애 핵심적인 역할을 하는 ATP 분자와 결합할 수 있는지를 조사했습니다. 다른 분자를 이동시키거나 새로운 형태로의 변환을 촉매하는 것과 같은 모든 유용한 작업을 하는 단백질은 ATP를 에너지 원으로 사용합니다. 따라서 ATP 결합체는 세포의 중요한 구성요소가 됩니다. 쇼스택과 키피는 화학적 방법으로 80개의 아미노산을 임의의 순서로 연결한 후, 이들의 모든 변이를 만들어 그 변이들이 우연히 ATP 와 결합할 수 있는지를 조사했습니다. 이들이 조사한 단백질의 수는 이 80개의 아미노산으로 만들 수 있는 전체 단백질 종류에 비하면 매우 작은 값입니다. 그러나 그들이 만든 6조개의 단백질 중에 그들은 4개의 ATP 결합체를 발견했습니다. 이 수는 많지 않아 보입니다. 그러나 전체 순서공간에서 극히 작은 이 부분 서열공간에 이미 4개의 해답이 있다는 것은, 10^93에 이르는 전체 서열 공간 안에는 무수히 많은 수의 해답이 있음을 의미합니다.
와그너와 그의 동료는 이런 “진화가능한(evolvable)” (그들은 견고한(robust)이라 불렀습니다) 구조가 생물학적 복잡성의 세상에서 공통으로 발견되는 특성임을 보였습니다. 2006년 그와 그의 동료 후안 로드리게즈는 박테리아 E. 콜리의 대사 네트웍 연구를 시작했습니다. E. 콜리는 모든 박테리아 중 가장 잘 연구된 박테리아이며 E. 콜리의 생화학적 대사경로는 잘 알려져 있습니다. E. 콜리는 포도당을 흡수해 이로부터 자신이 필요로 하는 60여개의 기본 분자들을 만듭니다. 그러나 대사 과정 중 하나가 바뀐다면 어떤 일이 벌어질까요? E. 콜리의 “유전자 이웃”이 포도당만으로 살아남을 수 있을까요? 가능한 E. 콜리의 유전자 이웃의 수는 매우 많지만, 와그너와 로드리게스는 그 중 1,000 개 이상의 이웃을 조사했고 그 중 수백개가 포도당만으로 살 수 있음을 보였습니다. 이는 E. 콜리의 대사 네트웍이 포도당에 맞도록 미세조정된 것이 아니며, 유전자 변이가 일어나더라도 역시 잘 작동한다는 것을 의미합니다. 연구진은 대사 공간을 확장해 이웃의 이웃들 역시 포도당을 소화할 수 있는지 조사했습니다. 그들은 포도당 소화가 가능한 대사 네트웍을 대사 공간을 따라 진행한 끝에 E. 콜리와는 단 20%의 반응만을 공유하는 가상의 미생물에 도달할 수 있었습니다. 이 경우에도 해답의 수는 무수히 많았고, 그 해답들은 가능 공간에서 중립 돌연변이들을 통해 도달가능했습니다. 진화는 수많은 미생물을 이 복잡한 도서관으로 보내 좋은 해답을 찾는 것으로 생명의 문제를 풀었던 것입니다. 와그너는 “대사 공간”에서 이런 예상치 못한 공간구조를 발견했을 때 “황홀한 기분이 들었다” 고 말합니다.
유전자 회로에도 같은 이야기가 있습니다. 와그너는 물리학자 올리비어 마틴과 가상의 유전자 회로 통제 네트웍을 조사했습니다. 그들은 구성요소의 활성화를 서로 통제하지만 표현형은 같은 패턴을 가지는 네트웍을 조사했습니다. 예를 들어, 유전자 A, B, C 로 이루어진 네트웍이 있다면, 유전자 A 가 유전자 B 를 억제할 수도 있고, 유전자 C 가 유전자 B 를 억제할 수도 있는 것입니다. 열 개 정도의 유전자만을 고려해도 가능한 네트웍의 수는 기하급수적으로 늘어나며, 연구자들은 오직 일부만을 확인할 수 있습니다. 그러나 결과는 분명했습니다. 유전자 회로는 일반적으로 같은 표현형을 가지는 수십에서 수백개의 이웃을 가졌으며, 또한 구성 요소가 90% 까지 다른 회로 중에도 같은 표현형을 가지는 회로가 존재했습니다.
이는 유전자 회로가 가진 묘한 특징인 ‘견고함’을 설명해 줍니다. 1990년대 후반, 스탠포드의 연구진은 맥주 효모의 유전자에서 서로 다른 유전자 하나를 제거해 6천여개의 변종을 만들었습니다. 그러나 이들 대부분은 원래 효모처럼 잘 번성했습니다. 다른 유기체들도 이런 특징을 가지고 있습니다. 각 유기체의 상당수 유전자는 제거해도 큰 효과가 나타나지 않습니다. 이는 다수의 유사한 유전자 회로가 원래의 유전자 회로와 같은 일을 한다는 것을 알면 놀라운 일이 아닙니다. 이렇게 말할 수 있겠지요. 곧, 견고함은 혁신과 보완적인 관계라고 말입니다. 진화를 통해 새로운 형태와 특징을 만들어 내기 위해서는 같은 성능을 내는 다른 대안들이 많아야 하므로, 결국 작은 변이에 대해 그 시스템은 ‘견고’해야 합니다. 이를 이해함으로써 우리는 과도한 유전자 결정론을 피할 수 있습니다. 유전자의 표현형은 유전자 하나가 아니라 유전자들이 만드는 네트웍에 보다 의존합니다.
스페인 바로셀로나 유전학 통제 연구소의 요하네스 예거는 와그너의 작업이 “실로 혁신적이며 중요”하다고 말합니다. 그는 초파리의 성장을 제어하는 유전자 네트웍의 진화 발달 생물학에 이들의 아이디어를 테스트했습니다. “지금까지 우리 실험 결과와 그의 주장이 일치합니다.” 그들은 초파리 기관의 발달 순서를 바꾸는 유전자 네트웍을 만드는 것이 생각보다 쉽다는 것을 발견했습니다. 곧, 전체 유전자를 바꿀 필요 없이 몇 가지 통제 기제를 조작하는 것만으로 그것은 가능했습니다. “이는 여러 초파리 종이 보이는 다양한 표현형을 설명합니다. 모든 종류의 패턴이 같은 종류의 유전자들로 만들어질 수 있습니다.”
진화는 자연의 법칙?
와그너의 발견은 생명의 한 특성을 보여주며, 이는 생명체를 만드는 진화과정 보다 더욱 심오한 것입니다. 그들은 진화가 일어나는 공간의 특성을 밝혔으며, 진화가 가능한 이유가 그 공간이 매우 특별한 구조를 가지고 있기 때문임을 보였습니다. 그 구조란, 유전자, 대사, 단백질 혹은 염기의 서열이 기능적으로 유사한 조합으로 연결된 거대한 거미줄로 다차원 공간을 채우고 있으며 각각은 또한 수많은 이웃들과 함께 엮여 있다는 것입니다.
어떤 이는 생명체가 탄생할 수 있었던 원인을 아미노산이나 설탕, 등 생명체를 구성하는 화학적 재료의 탄생이라 주장할 수 있습니다. 그러나 지금 이 시점에서 화학적으로 적당한 재료가 만들어진 것은 행복한 우연 정도의 상대적으로 쉬운 문제로 보입니다. 어려운 부분은 어떻게 이런 재료들로 가득찬 수프 상태에서 처음 진화가 시작되었나 하는 것입니다. 만루비아는 이 최초의 창조적 단계가 중립(혹은 아-중립) 네트웍의 풍부하고 밀접한 엮임의 결과라고 생각합니다. 이는 아직 생명의 역할을 하지 못하는 RNA 순서쌍들이라 하더라도 우연한 상황에서 어떤 유용한 기능을 하게 될 가능성이 충분히 크기 때문입니다. “어떤 면에서는 표현형이 서열 공간에서 충분한 부피를 차지하기만 하면, 특별한 기능을 공짜로 얻게 된다고 말할 수 있습니다.” 그녀는 컴퓨터 시뮬레이션으로 이를 만족하는 RNA 서열이 귀하지 않음을 보였습니다. “따라서 진화의 과정이 아직 시작되지 않은 상황에서, 진화 과정의 씨앗이 될 수 있는 충분히 많은 수의 해답들이 준비된 것입니다.” 특히, 어떤 서열은 자신을 복제할 수 있었을 가능성이 큽니다. 그리고 그 때부터 진화가 시작된 것입니다. “자연 선택은 어중간한 해답을 충분히 효율적인 해답으로 빠르게 변화시킵니다.”
이런 생체분자의 조합이 만든 특별한 공간 구조는 자연을 기존의 안정된 상태에서 작은 변화만을 보는 정도로 국한시키는 대신 보다 대담하고 창조적인 혁신을 시도하게 만들었습니다. 유기체는 적합도의 손실 없이도 중립 돌연변이의 거미줄을 따라 무작위로 돌아다닐 수 있었고, 같은 표현형이면서도 전혀 다른 이웃들을 가질 수 있었습니다. 그리고 환경이 변했을 때 이에 맞는 혁신적인 해답을 내놓을 수 있었습니다. 중립적인 돌연변이위의 표류는 유기체로 하여금 그들이 시작점에서 출발했다면 도달하지 못했을 적응적 변이를 가능하게 만들었습니다.
진화에 있어 이러한 무작위 표류의 역할은 오래 전부터 알려져 있었지만 최근 진화생물학자 존 타일러 보너는 이 무작위 표류가, 특히 다양한 형태와 구조를 가지는 작은 유기체 – 규조나 방산충과 같은 해양 미세 유기물 – 에 있어서는, 이들이 반드시 최적의 적응상태로 미세조절되어 있지 않다는 점에서 생각보다 더 중요한 것일수 있다고 주장하고 있습니다. 이 미생물들의 창조성과 예술성은 생물학자들을 놀라게 했고 예술가들에게 영감을 주었습니다. 이제 우리는 이들이 보이는 다양성과 독창성의 원인을 이해하게 된 듯 합니다.
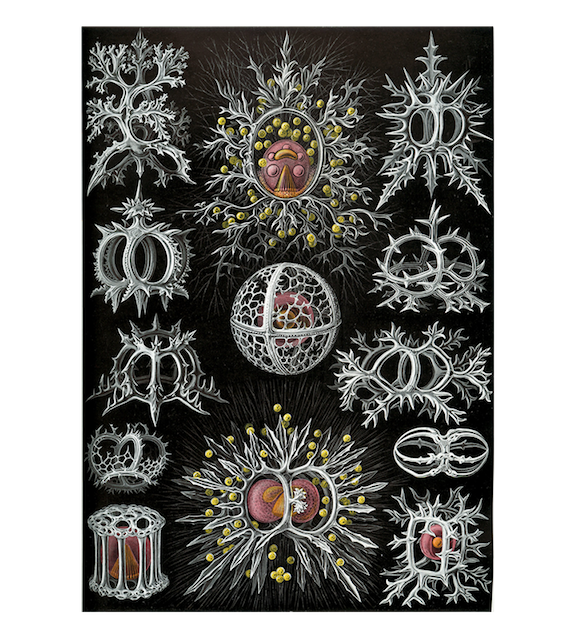
19세기 독일의 생물학자 에른스트 해켈이 그린 다양한 방산충
특히 와그너와 다른 이들의 업적은 진화에 있어 오랜 논쟁의 주제였던 집단의 적응과 개체의 적응 사이의 충돌 문제에 대한 답을 제시합니다. 어떤 박테리아들은 개체의 입장에서 보기에는 과한 정도의 돌연변이를 겪습니다. 만약 대부분의 돌연변이가 적응성을 낮춘다면, 이는 개체의 입장에서는 전혀 ‘현명한’ 일이 아닙니다. 그러나 단순하게 말하면, 집단의 입장에서는 새로운 환경에 적응할 수 있는 더 많은 선택지를 가지는 것이므로 이러한 개체의 과한 돌연변이가 이로운 특성이 됩니다. 하지만 와그너가 보인 것처럼, 견고한 네트웍이 중립적인 돌연변이의 확률을 높인다면, 바이러스와 같은 과도한 돌연변이도 개체에는 그렇게 나쁘지 않으면서 집단에게는 유리한 성질이 됩니다.
아직도 질문은 남아 있습니다. ‘왜 진화의 선택공간은 이렇게 견고한 구조를 가지게 되었나?’ 하는 것입니다. “우리는 왜 유전형 네트웍이 그렇게 얽혀있는지 알지 못합니다.” 와그너는 인정합니다. 단백질 진화에 대한 전문가인 시애틀 프레드 허친슨 암 연구소의 제시 블룸은 이 질문이 원인과 결과를 혼동한 질문이라고 생각합니다. “만약 진화가 이런 성질을 만족할 때에만 가능하다면, 진화가 이루어진 뒤에는 모두가 그런 성질을 가진 것으로 보이겠지요.” 그러나 그 역시 이를 증명하는 것은 매우 어려울 것임을 인정합니다.
어쩌면 이 문제는 생물학의 수준을 넘는 문제일지 모릅니다. 와그너의 연구실에 포닥으로 있었고 지금은 인도 마드라스 공대에 있는 카르틱 라만은 유전자가 아니라 이진 논리 기능을 가진 전기 회로의 구성요소들로 이루어진 회로에서 이와 같은 기능적 동등성이 있는지를 연구하고 있습니다. 16개의 구성요소를 가지고 특정한 논리적 기능을 가진 이 회로에서도 라만은 유전자 회로와 같은 진화가능한 구조가 있음을 발견했습니다. 하지만 놀라운 점은, 이러한 특성은 회로가 충분히 복잡할 때에만 나타났습니다. 회로의 구성요소가 너무 작아지면, 작은 변화로도 기능은 파괴되었습니다. “더 복잡할 수록, 변화에 대해 더 강했습니다.” 이 발견은 전기회로 디자인에도 다윈의 진화론을 적용할 수 있다는 것을 보였을 뿐 아니라 진화가능성, 곧, 창조성과 혁신성이 생물학에서만이 아니라 복잡한 네트웍의 기본적인 특징일 수 있음을 암시합니다.
만루비아 역시 복잡성이 핵심이라는 사실에 동의합니다. “효율적인 표류능력은 유전자 공간이 고차원일때만 가능하다는 것은 명백해 보입니다.” 이는 차원이 높을 수록 진행할 수 있는 방향이 많으며 따라서 더 많은 선택지를 가지기 때문입니다. “서열에 있어 가능한 이웃의 수가 증가할수록 이웃 중 원래의 서열과 동일한 기능을 발휘할 이웃이 존재할 확률 역시 증가합니다.” 그녀는 이로써 내릴 수 있는 한 가지 결론으로, 적어도 변화하는 환경에서 거주하는 유기체는 적응적으로 더 큰 게놈을 선호할 가능성이 있다고 말합니다. 더 많은 염색체를 유지하고 복사해야 하는 비용을 지불하는 대신 더 견고한 게놈을 가질 수 있게 되기 때문입니다.
이 아이디어들은 진화가능성과 혁신가능성이 생명의 특성만이 아니라 정보 자체의 특성 일 가능성을 암시합니다. 이는 슈스터의 한 때 동료였던 노벨 화학상 수상자인 만프레드 아이겐이 지지하는 생각입니다. 그는 다윈의 진화가 생물학에만 적용되는 원리가 아니며, “물리 법칙”으로 복잡한 시스템에서 정보가 구성되면서 필연적으로 발생하는 결과라고 주장합니다. 만약 그가 옳다면, 생명의 탄생은 환상적인 우연이 아니라 수학적으로 필연적인 사건이었을 것입니다.
(노틸러스)